植物似乎吸收了人工设计的针对特定基因的外源RNA,然后激活RNA干扰(RNAi)机制。然而,除了表明寄生植物与其宿主之间发生小RNA交换的证据外,植物是否使用RNA本身作为植物与植物交流中的信号分子尚不清楚。该研究发现植物RNA可以从一株传递到另一株并发挥功能。

小RNA(sRNA)是21到24个核苷酸的非编码分子,可影响植物的各种发育和生理过程。根据它们的生物发生和作用方式,有两组sRNA。Micro-RNA(miRNA)属于第一组,它包括由特定基因编码的sRNA,产生单链、自互补、非编码RNA,形成发夹结构,随后由Dicer样蛋白(DCL)加工以产生一个21到22个核苷酸的成熟miRNA。第二组包括小干扰RNA(siRNA),它是由依赖于RNA的RNA聚合酶6(RDR6)合成的双链RNA分子的DCL依赖性切割产生的。单链sRNA被加载到属于RNA诱导沉默复合物的ARGONAUTE(AGO)蛋白上。miRNA主要加载到AGO1蛋白上。RNA诱导的沉默复合物以序列同源性依赖的方式利用sRNA来识别mRNA靶标,这些靶标被切割并最终降解。
在植物内,sRNAs在局部起作用,但也可以从一个细胞移动到另一个细胞,大概是通过胞间连丝,并通过脉管系统进行长距离的系统运输。由sRNA触发的基因沉默最终可以在整个植物中传播。这些sRNA如何在植物内移动仍不清楚。miR399是响应磷酸盐饥饿而产生的,存在于几种植物的韧皮部汁液中,并且已被证明是韧皮部可移动的microRNA。在缺乏磷酸盐的植物中,miR399的表达增加,导致miR399易位到根系统,在那里它影响其目标的表达,即调节磷酸盐转运蛋白PHO1降解的E2结合酶PHO2。因此,miR399易位到缺磷植物的根系会导致PHO1的激活和磷酸盐吸收增加。
另一个经过充分研究的被认为是韧皮部可移动的miRNA是miR156。miR156通过抑制SPL转录因子来调节多种发育性状。在miR156/SPL模块调控的过程中,值得一提的是拟南芥中的幼年到成年的转变。由于幼年期延长,miR156的过表达的植株开花极晚。
有趣的是,sRNA不仅是植物内的移动信号和调节分子,而且还在植物和相互作用的生物体(包括病原体)之间移动,以诱导基因沉默。这种现象被称为跨界/生物体RNA干扰(RNAi)。为了响应大丽轮枝菌的感染,棉花植物显示出miR166和miR159的增加,并将两者输出到真菌菌丝以沉默对真菌毒力至关重要的两个轮枝菌基因。灰葡萄孢石竹变种将其sRNA传递到植物细胞中以沉默宿主免疫基因,宿主拟南芥细胞分泌外泌体样细胞外囊泡将植物sRNA传递到B.cinerea中。外源RNA可能被它们在叶表面接触的真菌细胞吸收,也可能先被植物细胞吸收,然后再转运到真菌细胞中。有趣的是,局部喷洒的双链RNA(dsRNA)也会抑制未处理的远端叶片的病原体毒力。这表明这些人工合成的dsRNAs在叶表面外用后在植物内系统性传播。
sRNA也在植物之间交换。菟丝子(Cuscutaspp.)是一种专性寄生植物,它使用吸器从寄主植物中获取水分和养分。菟丝子在寄生过程中积累了大量靶向拟南芥信使RNA的22核苷酸miRNA,导致mRNA裂解、次级siRNA产生和mRNA积累减少。这些结果表明来自菟丝子的miRNA作为宿主基因表达的跨物种调节剂,表明它们影响寄生植物的毒力。
如果来自环境的外源RNA被一些生物体吸收,它们可以诱导RNAi。这种现象被称为“环境RNAi”,已在线虫和昆虫中观察到,但未在植物中观察到。然而,针对昆虫发育基因的实验室合成dsRNA的局部应用会损害昆虫的生长。这些结果表明害虫内的基因表达通过摄取dsRNA或sRNA受到抑制;然而,目前尚不清楚昆虫吸收的RNA是在处理过的植物内部还是存在于其表面。
一些miRNA可以在植物中系统地发挥作用,表明它们作为移动信号分子发挥作用。然而,尚不清楚它们是否存在于环境中,它们是否被植物吸收,或者这是否最终导致接收植物中miRNA靶基因的转录后沉默。
该研究证明外源miRNA可以诱导转录后基因沉默(PTGS),并且相邻植物之间的miRNA转移可以影响基因表达。研究使用了两个miRNA模块,即miR156/SPL和miR399/PHO2,因为据报道它们都是在细胞间移动的,调节充分表征的植物过程并诱导次级siRNA的合成,潜在地放大目标mRNA的初始miRNA依赖性切割。
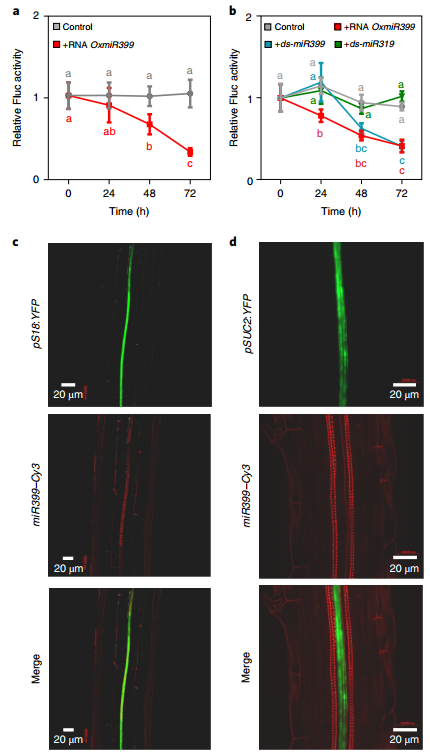
外源miRNA的易位和反应



